










Oyeleye I.S;1 Olofin A. T.;2 Oboh G.;3
1Department of Medical Biochemistry, Federal University of Technology, P.M.B. 704, Akure, Ondo State, Nigeria
2Functional Foods and Nutraceuticals Unit, Department of Biochemistry, Federal University of Technology, Akure, Ondo State, Nigeria
3Department of Pure and Applied Sciences, Precious Cornerstone University, Ibadan, Oyo State, Nigeria
4Functional Foods and Nutraceuticals Unit, Department of Biochemistry, Federal University of Technology, Akure, Ondo State, Nigeria
*Corresponding Author Email:goboh@futa.edu.ng…
Abstract
This study evaluates the effect of cooking on the neuronal activities of sweet, bird, and cayenne peppers. Three varieties of peppers (sweet, bird, and cayenne) were subjected to cooking for 20 min at 100°C. The raw and cooked peppers were analysed for their phenolic content, antioxidant activities, anti‐acetylcholinesterase, anti‐butyrylcholinesterase, and anti‐monoamine oxidase effects. The results indicated that cooked pepper had increased phenolic content, higher radical scavenging and metal‐chelating effects, but a reduced flavonoid level relative to raw pepper. Both raw and cooked samples inhibited AChE and BChE activities; however, the cooked samples showed greater inhibitory effects. A similar pattern was observed for MAO activity. Our findings suggest that cooking is effective in retaining the phenolic content of pepper, which may enhance its protective role against oxidative damage to neurons and cognitive function.
Keywords: Cooking effects, neuronal activities, antioxidant properties, phenolic content, cognitive function.
1. Introduction
Neurodegenerative diseases, such as Alzheimer’s disease (AD), are characterised by progressive and strategic loss of neuronal and cognitive functions (Kovacs, 2016). AD is characterised by a decline in acetylcholine (ACh) levels, progressive loss of cognitive function and oxidative stress, among others (Martin, 1999; Nieoullon, 2011). Cholinesterases (ChEs) are important biomarkers and are frequently targeted in the treatment of AD due to the rapid degradation of ACh in the synaptic cleft of neurons (Oboh et al., 2016). Low levels of ACh impair the cholinergic system and trigger cholinergic deficits, neuronal dysfunction, and cognitive decline (Schliebs & Arendt, 2011).
Monoamine oxidase, on the other hand, has also been identified as a target for the treatment of AD (Adefegha, Olasehinde, & Oboh, 2016). Monoamine oxidase (MAO) is involved in the oxidative degradation of biogenic amines such as dopamine, serotonin, and norepinephrine, thereby regulating the concentration of amine neurotransmitters as well as several amine drugs (McCabe et al., 2006). Previous reports have shown that MAO plays an important role in psychiatric conditions and neurological disorders (Bortolato & Shih, 2011; Adefegha, Olasehinde, & Oboh, 2016). Moreover, inhibition of MAO is an effective therapeutic strategy for the management of related mental disorders.
Pepper (Capsicum sp.) is a genus of flowering plants in the nightshade family (Solanaceae) that contains capsaicin as a pungent ingredient and has been reported to exhibit several pharmacological activities due to its richness in capsaicinoids, phenolic compounds, carotenoids, among others (Ruiz-Cruz et al., 2010; Alvarez et al., 2011). Pepper is consumed fresh, dried, smoked, pickled, or cooked for the preparation of sauces and as part of many different dishes. It is notable for its ability to break down lipids, thereby exhibiting protective effects against hypercholesterolaemia, obesity, and hypertension (Chopan & Littenberg, 2017). Previous reports from our laboratory revealed that hot pepper (Capsicum annuum, Tepin [CAT] and Capsicum chinense, Habanero) and bell pepper (Capsicum annuum var. grossum) protect the brain from toxicant-induced oxidative stress in rat brains and also inhibit β-secretase activities and β-amyloid aggregation (Oboh & Rocha, 2008; Ogunruku et al., 2016). Previous studies indicated that processing techniques could alter the health-promoting properties of plant foods (Lutz, Henriquez, & Escobar, 2011). In Nigeria, cooking is the most common processing method for peppers (Tiwari & Cummins, 2013). Interestingly, there is a paucity of information on the effect of cooking on the neuroactivities of pepper. Hence, this study evaluates the effects of cooking on the phenolic (total phenol and flavonoid) content, antioxidant activities, and enzyme (AChE, BChE, and MAO) inhibitory activities of three varieties of pepper (sweet pepper [Capsicum annuum L.], bird pepper [Capsicum frutescens L.], and cayenne pepper [Capsicum frutescens]).
2. Material and Methods
2.1 Sample Collection and Identification
Fresh sweet pepper [Capsicum annuum], bird pepper [Capsicum frutescens], and cayenne pepper [Capsicum frutescens] were purchased at Oja-Oba market in the Akure metropolis, south-west Nigeria. Authentication of the samples was carried out in the Department of Crop Soil and Pest Management, Federal University of Technology, Akure, Ondo State.
2.2 Preparation of Samples
All samples were washed with distilled water and allowed to drain. Each sample was then divided into two portions. One portion was cooked for 20 minutes at 100°C while the other was left uncooked (raw). Each portion was milled separately using an electrical laboratory blender. The milled portions were lyophilised using a freeze-dryer. Two grams of each sample portion were weighed and dissolved in 100 mL of distilled water, placed on an orbital shaker for 16 h, and then filtered through a muslin cloth. The filtrate was further centrifuged and filtered through Whatman No. 1 filter paper to obtain a clear filtrate, which was kept in the refrigerator and used for further analysis.
2.3 Chemicals and Reagents
Chemicals and reagents used were of analytical grade. Except where stated otherwise, all other chemicals and reagents were of analytical grade, and the water was glass-distilled.
2.4 Determination of Total Phenol and Flavonoid Content
The total phenol content was determined according to the method of Singleton et al. (1999), while the method of Meda et al. (2005) was used for the determination of total flavonoid content.
2.5 Antioxidant Capacity Assay
The reducing property of the extracts was determined by assessing their ability to reduce an FeCl₃ solution, as described by Oyaizu (1986). The total antioxidant capacity was determined based on the scavenging ability of the extracts against the 2,2′-azinobis(3-ethylbenzothiazoline-6-sulfonate) radical (ABTS*), according to the method described by Re et al. (1999). The free radical scavenging ability of the extracts against the 1,1-diphenyl-2-picrylhydrazyl (DPPH) radical was evaluated as described by Gyamfi, Yonamine, and Aniya (1999). The ability of the extracts to prevent Fe²⁺/H₂O₂-induced decomposition of deoxyribose was assessed using the method of Halliwell and Gutteridge (1981). The Fe²⁺ chelating ability of the extracts was determined using a modified method of Minotti and Aust (1987), with a slight modification by Puntel, Nogueira, and Rocha (2005). The lipid peroxidation assay was carried out using a modified method of Ohkawa, Ohishi, and Yagi (1979).
2.6 Acetylcholinesterase (AChE) and Butyrylcholinesterase (BChE) Inhibition Assays
Inhibition of AChE and BChE activity was assessed using a modified colorimetric method of Perry et al. (2000).
2.7 Monoamine Oxidase Inhibition Assay
The assay of MAO with different concentrations of the extracts was performed according to the methods of Green and Haughton (1961) and Turski et al. (1973), with slight modifications.
2.8 Data Analysis
The results of three replicate experiments were pooled and expressed as the mean ± standard deviation (SD). A one-way analysis of variance (ANOVA) followed by post hoc analysis using Duncan’s multiple range test was performed. Significance was accepted at P ≤ 0.05.
3. Results and Discussion
3.1 Effect of Cooking on Total Phenol and Flavonoid Content
The total phenol and flavonoid contents of the raw and cooked pepper extracts are presented in Table 1.

Among the raw peppers, RC had the highest total phenol content. There was no significant difference between the total phenolic content of bird (RB) and sweet (RS) pepper. Similarly, there was no significant difference (P > 0.05) in the total flavonoid contents of RB and RS, but these were significantly higher than that of RC. Moreover, cooked bird (CB), cayenne (CC) and sweet (CS) peppers had significantly (P < 0.05) higher total phenol content than raw bird (RB), cayenne (RC), and sweet (RS) peppers. However, RS and RB had significantly (P < 0.05) higher flavonoid contents than CS and CB extracts, respectively. Our findings revealed that cooking increased the total phenol content and reduced the flavonoid contents of the pepper extracts. Different processing techniques can influence the phytochemical composition of fruits and vegetables. Some reports indicated that cooking could reduce the phenolic content of peppers, while others report an increase. Hwang et al. (2012) reported that different cooking methods (boiling, roasting, and steaming) reduced the total phenol content of red pepper. Chuah et al. (2008) also reported a decline in the total phenol content of different varieties of peppers after cooking for 5 and 30 min. However, the report by Shotorbani et al. (2013) revealed that thermal treatment at 65°C increased the total phenol/flavonoid content of red pepper, but it decreased with an increase in exposure time beyond 60 min. There are indications that heat treatment may induce enzyme inactivation and the release of water-soluble constituents, which may influence the total phytochemical components of fruits and vegetables (Leong & Oey, 2012). The high phenolic contents of the cooked pepper observed in this study may be due to the release of bound phenols, which are liberated due to the disruption of cell membranes or structures upon heating (Kunyanga et al., 2012). Moreover, there are reports that heat treatment can improve the extraction of some phytochemicals (Howard et al., 1999; Leong & Oey, 2012). The levels of phytochemicals retained in vegetables after heat treatment depend on their stability (Leong & Oey, 2012). In this study, the observed low flavonoid contents in the cooked pepper extracts could be due to the degradation of some flavonoids during the heat process.
3.2 Effect of Cooking on the Antioxidant Activity of Raw and Cooked Pepper Extracts
The antioxidant properties of the samples were evaluated using the ferric-reducing antioxidant power (FRAP) assay and radical scavenging abilities (ABTS, DPPH, OH). As presented in Table 1, all the samples exhibited the capacity to reduce Fe³⁺ to Fe²⁺. Moreover, cooking increased the ferric-reducing ability of bird pepper. In contrast, cooking reduced the ferric-reducing capacity of cayenne pepper compared to its raw counterpart. However, there was no significant difference between the raw and cooked sweet pepper extracts.
The ability of the pepper extracts to decolourise the ABTS radical was evaluated. The capacity of the raw and cooked pepper extracts to scavenge the ABTS radical ranged from 109.80 to 227.45 mg/g Trolox equivalent antioxidant capacity (TEAC), as shown in Table 1.
Cooking increased the ABTS radical scavenging activity of sweet and cayenne pepper, but reduced that of bird pepper when compared to its raw extract. The highest ABTS radical scavenging activity was observed with RB, followed by RS. However, after cooking for 20 minutes, CS exhibited the highest ABTS radical scavenging activity compared to the other cooked peppers.
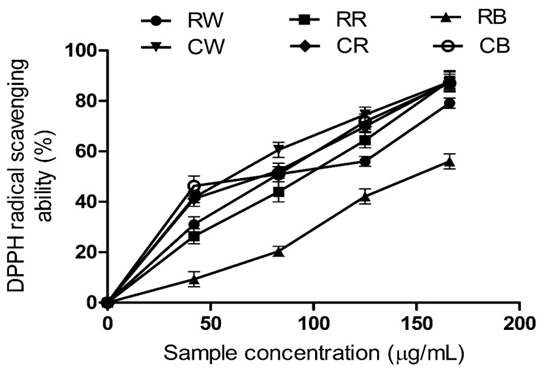
Figure 1: DPPH• Scavenging Ability of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper, and Cooked Cayenne Pepper
KEY: RW (Raw Ata wewe): Raw Bird pepper; CW (Cooked Ata wewe): Cooked Bird pepper; RR (Raw Ata rodo): Raw Sweet pepper; CR (Cooked Ata rodo): Cooked Sweet pepper; RB (Raw Bawa): Raw Cayenne pepper; CB (Cooked Bawa): Cooked Cayenne pepper
Table 2: IC₅₀ of OH•, DPPH• Scavenging and Fe²⁺ Chelating Abilities of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper and Cooked Cayenne Pepper (µg/ml)

Note: Values with the same superscript letter in the same column are not significantly different (P > 0.05).
The IC₅₀ values for the DPPH radical scavenging activities of the raw and cooked pepper extracts revealed that RB, RS, and RC exhibited significantly (P < 0.05) lower DPPH radical scavenging activities compared to their cooked counterparts (Table 2 and Figure 1). Cooking improved the DPPH radical scavenging of the studied pepper varieties. The observed higher radical scavenging activity of the cooked pepper extracts correlates with their high phenolic contents.
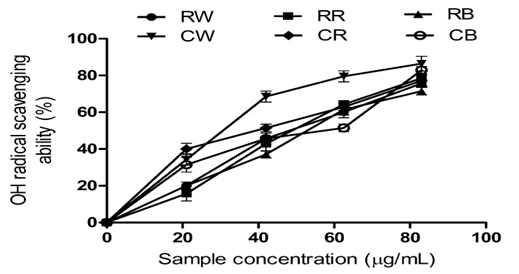
Figure 2: Hydroxyl Radical (OH•) Scavenging Abilities of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper, and Cooked Cayenne Pepper
KEY:RW (Raw Ata wewe): Raw Bird pepper; CW (Cooked Ata wewe): Cooked Bird pepper; RR (Raw Ata rodo): Raw Sweet pepper; CR (Cooked Ata rodo): Cooked Sweet pepper; RB (Raw Bawa): Raw Cayenne pepper; CB (Cooked Bawa): Cooked Cayenne pepper
Figure 2 depicts the hydroxyl (OH) radical scavenging activities of the pepper extracts. The IC₅₀ values revealed in Table 2 indicate no significant difference (P > 0.05) between raw bird pepper (RB) and raw sweet pepper (RS); however, both extracts exhibited higher OH radical scavenging activity compared to RC. After cooking for 20 min, the cooked pepper extracts exhibited higher OH radical scavenging activity than the raw extracts. Hydroxyl radicals are highly reactive and could cause damage to lipids, DNA, and proteins (Valko et al., 2007). The observed higher OH radical scavenging effect could prevent hydroxyl radical-induced oxidative damage to macromolecules within brain cells and contribute to their protective potential against lipid peroxidation.
Figure 3: Fe²⁺ Chelating Abilities of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper, and Cooked Cayenne Pepper.
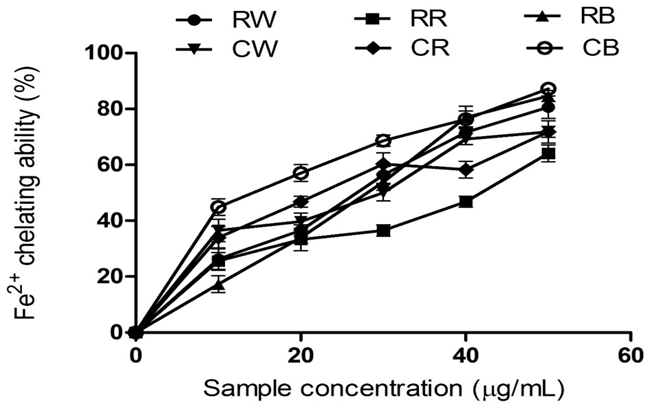
KEY: RW (Raw Ata wewe): Raw Bird pepper; CW (Cooked Ata wewe): Cooked Bird pepper; RR (Raw Ata rodo): Raw Sweet pepper; CR (Cooked Ata rodo): Cooked Sweet pepper; RB (Raw Bawa): Raw Cayenne pepper; CB (Cooked Bawa): Cooked Cayenne pepper
Figure 3 reveals the ability of the pepper extracts to chelate metal (Fe²⁺) ions. Fe²⁺ can catalyse one-electron transfer reactions that generate reactive oxygen species, such as the reactive OH, which is formed from H₂O₂ through the Fenton reaction (Valko et al., 2007). Iron also decomposes lipid peroxides, thereby generating peroxyl and alkoxyl radicals that are capable of initiating lipid peroxidation (Zago Verstraeten & Oteiza, 2000). In this study, cooking increased the metal-chelating ability of the pepper extracts, except for the bird pepper extracts, which showed no significant difference between the raw (RB) and cooked (CB) extracts. Moreover, the metal-chelating capacity of the pepper extracts could be linked to their phenolic content.
3.3 Cholinesterase Inhibitory Activity of Pepper Extracts
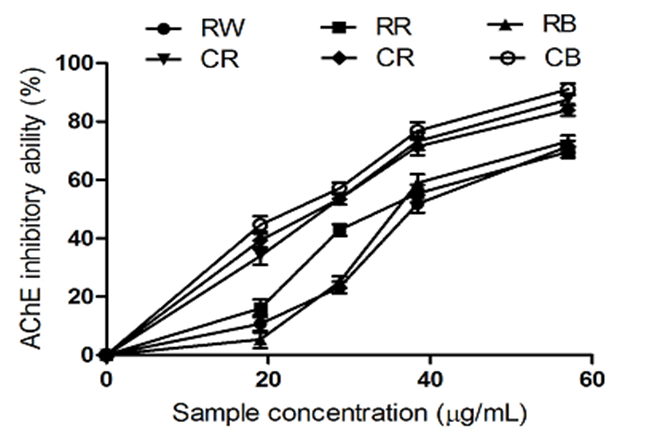
Figure 4: The Inhibitory Activity of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper, and Cooked Cayenne Pepper on Acetylcholinesterase
KEY: RW (Raw Ata wewe): Raw Bird pepper; CW (Cooked Ata wewe): Cooked Bird pepper;RR (Raw Ata rodo): Raw Sweet pepper; CR (Cooked Ata rodo): Cooked Sweet pepper; RB (Raw Bawa): Raw Cayenne pepper; CB (Cooked Bawa): Cooked Cayenne pepper
Table 3: IC₅₀ Values for Inhibition of Acetylcholinesterase, Butyrylcholinesterase and Monoamine Oxidase of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper and Cooked Cayenne Pepper (µg/ml)
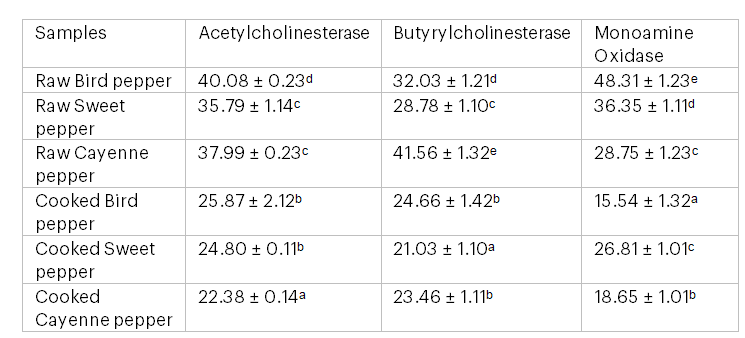
Note: Values with the same superscript letter in the same column are not significantly different (P > 0.05).
The AChE inhibitory activity of the pepper extracts is presented in Figure 4. The extracts inhibited AChE in a dose-dependent manner. According to the IC₅₀ values in Table 3, there was no significant difference between the AChE inhibitory activity of raw sweet pepper (RS) and raw cayenne pepper (RC); however, both exhibited higher inhibitory effects compared to raw bird pepper (RB). The AChE inhibitory activity of the cooked pepper extracts increased significantly compared to that of the raw extracts. AChE is an important target in the management of AD. An increase in AChE activity in AD patients reduces ACh levels—a major neurotransmitter responsible for the transmission of nerve impulses from one neuron to another (Oboh et al., 2017). Depletion of acetylcholine due to the upregulation of acetylcholinesterase activity causes cognitive deficits and impairments in learning functions (Olasehinde, Olaniran, & Okoh, 2017).
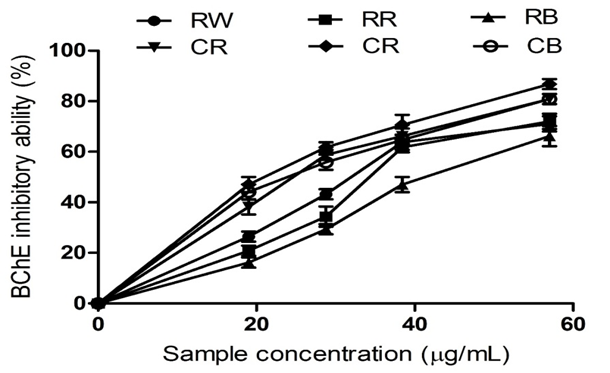
Figure 5: The Inhibitory Activity of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper, and Cooked Cayenne Pepper on Butyrylcholinesterase.S
KEY: RW (Raw Ata wewe): Raw Bird pepper; CW (Cooked Ata wewe): Cooked Bird pepper; RR (Raw Ata rodo): Raw Sweet pepper; CR (Cooked Ata rodo): Cooked Sweet pepper; RB (Raw Bawa): Raw Cayenne pepper; CB (Cooked Bawa): Cooked Cayenne pepper.
Inhibition of AChE increases ACh levels in the synaptic cleft and improves the transmission of signals between neurons. Our findings suggest that cooking improves the anti-AChE activity of pepper extracts. Similarly, the pepper extracts inhibited BChE activity, as depicted in Figure 5. RS presented a significantly higher inhibitory effect compared to RB and RC, as shown by their IC₅₀ values in Table 2. Cooking increased the inhibition of BChE activity. Although AChE plays a dominant role in a healthy brain, BChE also plays a minor role in the regulation of ACh levels. BChE activity increases progressively in AD patients, which may lead to a cholinergic deficit and cognitive dysfunction (Greig, Lahir, & Sambarmuti, 2002). The inhibitory effects of the pepper extracts on AChE and BChE may be linked to their phenolic and flavonoid content. However, the higher phenolic content of the cooked pepper extracts correlates with the observed higher anti-cholinesterase activity. Phenolic compounds have been proven to be potent inhibitors of AChE and BChE. Previous reports from our laboratory have shown that phenolic extracts containing gallic acid, chlorogenic acid, caffeic acid, catechin, epicatechin, rutin, and quercetin are potent inhibitors of AChE and BChE (Oboh et al., 2017).
3.4 Monoamine Oxidase Inhibitory Activity of Pepper Extracts
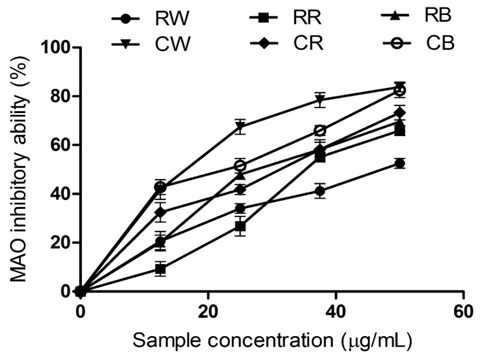
Figure 6: The Inhibitory Activity of Raw Bird Pepper, Raw Sweet Pepper, Raw Cayenne Pepper, Cooked Bird Pepper, Cooked Sweet Pepper, and Cooked Cayenne Pepper on Monoamine Oxidase
KEY: RW (Raw Ata wewe): Raw Bird pepper; CW (Cooked Ata wewe): Cooked Bird pepper; RR (Raw Ata rodo): Raw Sweet pepper; CR (Cooked Ata rodo): Cooked Sweet pepper; RB (Raw Bawa): Raw Cayenne pepper; CB (Cooked Bawa): Cooked Cayenne pepper
A decrease in the levels of monoamine neurotransmitters at major sites within the brain, due to an increase in monoamine oxidase (MAO) activity, is a major factor that leads to the onset of depression (Meyer et al., 2006). Figure 6 depicts the effects of the pepper extracts on MAO. The raw pepper extracts caused a decrease in MAO activity, but RC exhibited the highest MAO inhibitory effect compared to RS and RB, as revealed by their IC₅₀ values in Table 2. However, cooking resulted in a significant increase in the inhibition of MAO activity. CS exhibited a higher inhibitory effect compared to CB and CC. The observed increase in the enzyme inhibitory effect of the cooked pepper extracts could be attributed to the phenolic content. This result also suggests that heat treatment during cooking could improve the extractability of some phytochemical compounds or convert them into bioactive species with a potent capacity to inhibit MAO (Schweiggert, Schieber, & Carle, 2006). The inhibitory effect of the pepper extracts on monoamine oxidase could improve monoaminergic neurotransmission, enhance dopamine levels, and reduce behavioural disturbances. Furthermore, the observed inhibition by the pepper extracts may also prevent the generation of free radicals induced by the upregulation of MAO, thereby preventing neuronal damage in PD patients (Adefegha et al., 2016).
4. Conclusion
Cooking increased the total phenol content, antioxidant activity, anticholinesterase, and monoamine oxidase inhibitory activities of bird, sweet, and cayenne pepper. A loss of flavonoids was observed in the cooked pepper extracts. This finding reveals that cooking peppers is suitable for ensuring higher retention of phenolic compounds, which may promote cognitive function. The AChE, BChE, and MAO inhibitory activities of the extracts could be associated with their phenolic contents.
Conflict of Interest
The authors declare that they have no conflicts of interest related to this manuscript.
References
Adefegha, S. A., Oboh, G., & Olasehinde, T. A. (2016). Alkaloid extracts from shea butter and breadfruit as potential inhibitors of monoamine oxidase, cholinesterases, and lipid peroxidation in rats’ brain homogenates: A comparative study. Comparative Clinical Pathology, 25(6), 1213–1219.
Alvarez‐Parrilla, E., de la Rosa, L. A., Amarowicz, R., & Shahidi, F. (2011). Antioxidant activity of fresh and processed Jalapeño and Serrano peppers. Journal of Agricultural and Food Chemistry, 59(1), 163–173.
Bortolato, M., & Shih, J. C. (2011). Behavioral outcomes of monoamine oxidase deficiency: Preclinical and clinical evidence. International Review of Neurobiology, 100, 13–42.
Chopan, M., & Littenberg, B. (2017). The association of hot red chili pepper consumption and mortality: A large population-based cohort study. PLoS ONE, 12(1), e0169876.
Chuah, A. M., Lee, Y., Yamaguchi, T., Takamura, H., Yin, L., & Matoba, T. (2008). Effect of cooking on the antioxidant properties of coloured peppers. Food Chemistry, 111, 20–28.
Green, A. L., & Haughton, T. M. (1961). A colourimetric method for the estimation of monoamine oxidase. Biochemical Journal, 78, 172.
Greig, N. H., Lahiri, D. K., & Sambamurti, K. (2002). Butyrylcholinesterase: An important new target in Alzheimer’s disease therapy. International Journal of Psychogeriatrics, 14(Suppl 1), 77–91.
Gyamfi, M. A., Yonamine, M., & Aniya, Y. (1999). Free‐radical scavenging action of medicinal herbs from Ghana: Thonningia sanguinea on experimentally induced liver injuries. General Pharmacology, 32, 661–667.
Halliwell, B., & Gutteridge, J. M. C. (1981). Formation of a thiobarbituric‐acid‐reactive substance from deoxyribose in the presence of iron salts. FEBS Letters, 128, 347–352.
Howard, L. A., Wong, A. D., Perry, A. K., & Klein, B. P. (1999). Beta‐carotene and ascorbic acid retention in fresh and processed vegetables. Journal of Food Science, 64(5), 929–936.
Hwang, I. G., Shin, Y. J., Lee, S., Lee, J., & Yoo, S. M. (2012). Effects of different cooking methods on the antioxidant properties of red pepper (Capsicum annuum L.). Preventive Nutrition and Food Science, 17(4), 286–292.
Kovacs, G. G. (2016). Molecular pathological classification of neurodegenerative diseases: Turning towards precision medicine. International Journal of Molecular Sciences, 17, 189.
Kunyanga, C. N., Imungia, J. K., Okotha, M. W., Biesalski, H. K., & Vadivel, V. (2012). Total phenolic content, antioxidant and antidiabetic properties of methanolic extract of raw and traditionally processed Kenyan indigenous food ingredients. LWT – Food Science and Technology, 45, 269–276.
Leong, Y., & Oey, I. (2012). Effects of processing on anthocyanins, carotenoids and vitamin C in summer fruits and vegetables. Food Chemistry, 133, 1577–1587.
Lutz, M., Henríquez, C., & Escobar, M. (2011). Chemical composition and antioxidant properties of mature and baby artichokes (Cynara scolymus L.), raw and cooked. Journal of Food Composition and Analysis, 24, 49–54.
Martin, J. B. (1999). Molecular basis of the neurodegenerative disorders. New England Journal of Medicine, 340, 1970–1980.
Meda, A., Lamien, C. E., Romito, M., Millogo, J., & Nacoulma, O. G. (2005). Determination of the total phenolic, flavonoid and proline contents in Burkina Faso honey, as well as their radical scavenging activity. Food Chemistry, 91, 571–577.
Meyer, J. H., Ginovart, N., Boovariwala, A., Sagrati, S., Hussey, D., Garcia, A., Young, T., Praschaik-Rieder, N., Wilson, A. A., & Houle, S. (2006). Elevated monoamine oxidase A levels in the brain: An explanation for the monoamine imbalance of major depression. Archives of General Psychiatry, 63, 1209–1216.
Minotti, G., & Aust, S. D. (1987). An investigation into the mechanism of citrate Fe²⁺-dependent lipid peroxidation. Free Radical Biology and Medicine, 3, 379–387.
Nieoullon, A. (2011). Neurodegenerative diseases and neuroprotection: Current views and prospects. Journal of Applied Biomedicine, 9(4), 173–183.
Oboh, G., Nwanna, E. E., Oyeleye, S. I., Olasehinde, T. A., Ogunsuyi, O. B., & Boligon, A. A. (2017). In vitro neuroprotective potentials of aqueous and methanol extracts from Heinsia crinita leaves. Food Science and Human Wellness, 5, 95–102.
Oboh, G., Ogunruku, O. O., Oyeleye, S. I., Olasehinde, T. A., Ademosun, A. O., & Boligon, A. A. (2016). Phenolic extracts from Clerodendrum volubile leaves inhibit cholinergic and monoaminergic enzymes relevant to the management of some neurodegenerative diseases. Journal of Dietary Supplements, 14(3), 358–371.
Oboh, G., & Rocha, J. B. (2008). Hot pepper (Capsicum spp.) protects brain from sodium nitroprusside- and quinolinic acid-induced oxidative stress in vitro. Journal of Medicinal Food, 11(2), 349–355.
Ohkawa, H., Ohishi, N., & Yagi, K. (1979). Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Analytical Biochemistry, 95, 351–358.
Olasehinde, T. A., Olaniran, A. O., & Okoh, A. I. (2017). Therapeutic potentials of microalgae in the treatment of Alzheimer’s disease. Molecules, 22(3), 480.
Oyaizu, M. (1986). Studies on products of browning reaction: Antioxidative activity of products of browning reaction prepared from glucosamine. Japan Journal of Nutrition, 44, 307–315.
Perry, N. S., Houghton, P. J., Theobald, A., Jenner, P., & Perry, E. (2000). In vitro activity of Salvia lavandulaefolia (Spanish sage) relevant to treatment of Alzheimer’s disease. Journal of Pharmacology, 52, 895–902.
Puntel, R. L., Nogueira, C. W., & Rocha, J. B. T. (2005). Krebs cycle intermediates modulate thiobarbituric acid reactive species (TBARS) production in rat brain in vitro. Neurochemical Research, 30, 225–235.
Re, R., Pellegrini, N., Proteggente, A., Pannala, A., Yang, M., & Rice-Evans, C. (1999). Antioxidant activity applying an improved ABTS radical cation decolourisation assay. Free Radical Biology and Medicine, 26, 1231–1237.
Ruiz-Cruz, S., Alvarez-Parrilla, E., de la Rosa, L. A., Martinez-Gonzalez, A. I., Ornelas-Paz, J. J., Mendoza-Wilson, A. M., & Gonzalez-Aguilar, G. A. (2010). Effect of different sanitizers on microbial, sensory and nutritional quality of fresh-cut Jalapeño peppers. American Journal of Agricultural and Biological Sciences, 5(3), 331–341.
Schliebs, R., & Arendt, T. (2011). The cholinergic system in aging and neuronal degeneration. Behavioural Brain Research, 221, 555–563.
Schweiggert, U., Schieber, A., & Carle, R. (2006). Effects of blanching and storage on capsaicinoid stability and peroxidase activity of hot chili peppers (Capsicum frutescens L.). Innovative Food Science & Emerging Technology, 7, 217–224.
Shotorbani, N. Y., Jamei, R., & Heidari, R. (2013). Antioxidant activities of two sweet pepper (Capsicum annuum L.) varieties phenolic extracts and the effects of thermal treatment. Avicenna Journal of Phytomedicine, 3, 25–34.
Singleton, V. L., Orthofer, R., & Lamuela-Raventós, R. M. (1998). Analysis of total phenols and other oxidation substrates and antioxidants by means of the Folin-Ciocalteu reagent. In R. M. Lamuela-Raventós (Ed.), Methods in Enzymology (Vol. 299, pp. 152–178). Academic Press.
Tiwari, U., & Cummins, E. (2013). Factors influencing levels of phytochemicals in selected fruit and vegetables during pre- and post-harvest food processing operations. Food Research International, 50(2), 497–506.
Turski, W., Turska, E., & Bellard, G. M. (1973). Modification of the spectrophotometric method of the determination of monoamine oxidase. Vopr. Med. Khim., 14, 211–220.
Valko, M., Leibfritz, D., Moncol, J., Cronin, M. M., Mazur, M., & Telser, J. (2007). Free radicals and antioxidants in normal physiological functions and human disease. International Journal of Biochemistry & Cell Biology, 39, 44–84.
Zago, M. P., Verstraeten, S. V., & Oteiza, P. I. (2000). Zinc in the prevention of Fe²⁺ initiated lipid and protein oxidation. Biological Research, 33(2), 143–150.
About this Article
Cite this Article
APA
Oyeleye I.S., Olofin A. T. & Oboh G. (2025). Cooking Enhances the Phenolic Content, Cholinesterase, and Monoamine Oxidase Inhibitory Activities of three Varieties of Pepper Fruits (Capsicum spp.). In Akinyele B.J., Kayode R. & Akinsemolu A.A. (Eds.), Microbes, Mentorship, and Beyond: A Festschrift in Honour of Professor F.A. Akinyosoye. SustainE
Chicago
Oyeleye I.S., Olofin A. T., and Oboh G. 2025. “Cooking Enhances the Phenolic Content, Cholinesterase, and Monoamine Oxidase Inhibitory Activities of three Varieties of Pepper Fruits (Capsicum spp.)” In Microbes, Mentorship, and Beyond: A Festschrift in Honour of Professor F.A. Akinyosoye, edited by Akinyele B.J., Kayode R. and Akinsemolu A.A., SustainE.
Received
15 November 2024
Accepted
10 January 2025
Published
4 February 2025
Corresponding Author Email: goboh@futa.edu.ng
Disclaimer: The opinions and statements expressed in this article are the authors’ sole responsibility and do not necessarily reflect the viewpoints of their affiliated organizations, the publisher, the hosted journal, the editors, or the reviewers. Furthermore, any product evaluated in this article or claims made by its manufacturer are not guaranteed or endorsed by the publisher.
Distributed under Creative Commons CC-BY 4.0
Share this article
Use the buttons below to share the article on desired platforms.




{kind=link}